David Cappaert · 8
Andrena chlorogaster, JF, dorsal, IAE |
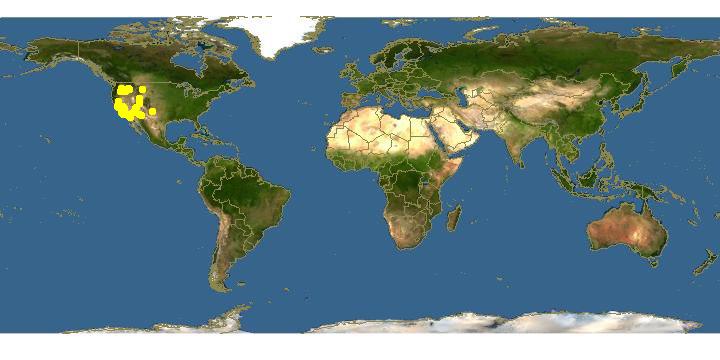
Click on map for details about points.
|
|
Overview |
Reprinted with permission from the University of Nebraska State Museum from:
Ribble, D. W. 1968. Revisions of two subgenera of Andrena, Micrandrena Ashmead and Derandrena new subgenus (Hymenoptera: Apoidea). Bulletin of the University of Nebraska State Museum 8: 1-124.
Please report text errors to: leah at discoverlife dot org.
In his original description of nitidicornis (one specimen from the northern coast of California) Cockerell postulated that it may be the male of chlorogaster described from Oregon. Andrena nitidicornis has been considered a distinct species because it differs in many respects from the "typical" chlorogaster. Considerable time has been spent during this study trying to separate chlorogaster from nitidicornis, but characters have not been found that did not often break down. Even when using several characters, material could not be placed with certainty. Evidence most strongly indicates that nitidicornis is merely a geographic variation of chlorogaster. Andrena nitidicornis specimens are found in the mountains of California while the typical chlorogaster are found in the Sacramento and San Joaquin Valleys. However, there are many exceptions to these ranges and mixing of the two is indicated, so much so that there seems to be little restriction in gene flow. In southern California where most specimens are intermediates and where populations in close geographic proximity may be noticeably different (see Variation for more details), the relation is more complex. Timberlake (1951) was led to recognize three of these populations as distinct species or subspecies as well as nitidicornis and chlorogaster. However, he had limited material which indicated several distinct forms rather than one highly variable population.
Andrena chlorogaster nesiotes is known from a single male from Santa Catalina Island. Timberlake (1951) described nesiotes on the basis of genitalia, antenna length and color, and facial hair color. The condition of these characters exhibited by nesiotes falls within the range of mainland variation of chlorogaster, i.e., males collected near King City, California, are nearly identical to nesiotes in all characters listed above.
The type series of chlorogaster gavilanica contained a single male (holotype) and several females. Timberlake (1951) was not able to separate the females from "typical" chlorogaster. The male from The Gavilan (near Lake Mathews, Riverside County, California) was distinguished on the basis of flagellar color, facial hair color and genitalia. The first two characters are highly variable throughout the range of chlorogaster and represent individual variation. The male genitalia of gavilanica are nearly intermediate between chlorogaster and what has been known as nitidicornis and there is no reason to recognize this genitalic condition as a subspecific character to do so would lead to naming a number of populations which are really just states within the continuous variation of chlorogaster.
Andrena radialis was described from a long series of males and females taken within a 50 mile radius of Riverside. Timberlake (1951) separated the females on the basis of the whitish to veal hair color, but even this character, he states, may not always be distinctive. Indeed, females with whitish foveae occur throughout the range of chlorogaster. Two additional characters are used in his key: more curled scopal hairs and ferruginous pterostigma and wing veins. Both of these characters are too variable throughout the range of chlorogaster to be of value in distinguishing radialis. Several characters were used to describe the male of radialis, but none of these characters would seem to warrant recognizing radialis. For example, specimens from northern Utah fit the description of radialis except in the genitalic characters. The genitalia of radialis are much like those of gavilanica and represent a slightly different intermediate condition between chlorogaster and what has been previously known as nitidicornis (see Variation).
The only known female from Santa Catalina Island was described as Andrena catalinica. This specimen is very fresh looking and agrees most closely with some specimens collected in the Berkeley, California, area. The only difference between the holotype and these mainland specimens is that the type has some chocolate brown hairs on the dorsum of the thorax and along the inner eye margins, while the same hairs on the Berkeley specimens are yellowish. Different hair colors are thought to be largely a variation due to age; dark hairs in time fade to white. Other species of Micrandrena show this same type of hair color variation. The male holotype of nesiotes from the same island, collected 17 days later in another year, has all white hairs. For these reasons and those listed under nesiotes an island race is not recognized. The holotype and only known specimen of subtilicornis was not located. The description and drawings in the original publication (Viereck, 1926) indicate subtilicornis belongs here. Viereck did not mention the clypeus color but implied it was yellow when he placed subtilicornis near personata. Cockerell said the face was dark (1929, 1932) but admitted he had not seen the type (Cockerell, 1932). Cockerell (1929) thought subtilicornis to be, perhaps, nearest to timberlakei in the California fauna.
The males of chlorogaster can be distinguished by their long, shiny antennae and yellow clypeus without lateral hyaline spots. The yellow maculation is often much reduced so that the clypeus is mostly dark. The females have a moderately wide hind tibia with scopal hairs of medium length and the second or third segments of the flagellum are subequal to the fourth. In both sexes the integument gives off weak metallic reflections or is without these reflections; the integument of microchlora has strong metallic reflections. The sexes of chlorogaster have a large pterostigma and usually the first transverse cubital vein ends very near the pterostigma.
FEMALE. MEASUREMENTS AND RATIOS. Length, 6.6 mm; width of mesothorax, 1.9 mm; wing length from tegula, 5.3 mm; facial length/width, 0.97; foveal length/width, 3.0.
INTEGUMENTAL COLOR. Black to dark brown with slight greenish-gold reflections except: flagellum dark brown below at tip; clypeus with purplish reflections; veins and pterostigma dark amber; terga and sterna with amber apices; color otherwise as in piperi.
STRUCTURE. Sculpturing in general finer over entire body than in piperi. Head otherwise as in piperi except: flagellar segment 1 slightly shorter than 2 and 3 together; segment 2 slightly shorter than 3; segments 3 and 4 equal in length; middle segments about as wide as long; eye about four times as long as broad in anterior view; malar space about seven times as wide as long; maxillary palpus with segments in ratio of about 1.1:1.1:1.0:1.0:1.0:1.3; labial palpus with segments in ratio of about 2.0:1.0:1.0:1.4; labral process one-third as wide and two-thirds as long as entire labrum, base expanded, apex rounded; genal area in lateral view slightly wider than eye; surface between ocelli moderately rough; surface above fovea not smoother, not more shiny than surface between ocelli.
Mesosoma as in piperi except enclosure of propodeum with rather even reticular shagreening turning into fine rugulae anteriorly.
Metasoma as in piperi.
VESTITURE. Hairs as in piperi except: dorsum of thorax with relatively short, plumose hairs, moss-like appearance less noticeable than in piperi; terga 1 and 2 with few minute, mostly simple hairs, except distally and laterally; terga 3 and 4 hairs longer, very numerous, most hairs distinctly branched; apical fasciae on terga 2-4, interrupted medially on 2 and 3, complete on 4; tibial scopal hairs moderately long, simple, posterior- and anterior-most hairs may have a few branches, hairs along anterior margin curved slightly posteriorly, hairs somewhat overlapping, most hairs tend to be orientated posteriorly, surface of tibia partly obscured by hairs.
MALE. MEASUREMENTS AND RATIOS. Length, 5.9 mm; width of mesothorax, 1.6 mm; wing length from tegula, 4.9; facial length/width, 0.96; flagellar segment 1/segment 2, 0.65.
INTEGUMENTAL COLOR. Black with slight bluish reflections except:
clypeus yellow without lateral hyaline spots; flagellum yellow-brown below; veins dark amber; pterostigma black; color otherwise as in piperi.
STRUCTURE. Sculpturing in general finer over entire body than in piperi. Head otherwise as in piperi except: antenna reaching abdomen; scape reaching to within two ocellar distances of median ocellus, much shorter than flagellar segments 1-3; segment 1 two-thirds as long as 2 or 3; middle segments one and one-half times as long as broad; malar space about six times as wide as long; maxillary palpus with segments in ratio of about 1.1:1.3:1.0:1.1:1.1:1.3; labial palpus with segments in ratio of about 1.8:1.2:1.0:1.4; clypeus about one and one-half times as broad as long; genal area slightly wider than eye.
Mesosoma as in chlorogaster female except: metanotum more coarsely sculptured than mesoscutum; posterior and dorsolateral areas not raised above enclosure; tibia unmodified.
Exposed metasoma as in piperi.
Sternum 7 truncate at apex, producing squared process, corners of process with few, long, branched setae (fig. 47). Sternum 8 (fig. 48) much like piperi, except basal plate different in outline. Gonocoxite bent downward, apex expanded in lateral view, dorsolateral part with many weakly branched setae; dorsal lobes short. Penis valve bent downward, apex very broadly rounded in lateral view, medially expanded; subapical lamellae greatly humped, extending posteriorly past gonocoxite (figs. 41-43).
VESTITURE. Hairs as in piperi except: head with few dark hairs along eye; hairs on abdominal terga sparser and less plumose.
VARIATION. Within California, chlorogaster exhibits considerable geographic variation. If we exclude southern California (Ventura County and southward) a trend can be seen. Specimens from the Sacramento and San Joaquin valleys (hereafter referred to as the valley condition) tend to show one group of characters while specimens from the mountains surrounding these valleys (Coast Range and Sierra Nevada) (hereafter referred to as the montane condition) tend to show another group of characters. Although there are exceptions to this distribution of characters and numerous examples of intermediate specimens are known, the overall tendencies are clear.
In some females the hairs on the medial area of abdominal terga 3 and 4 are numerous, branched and of medium length. These hairs partly obscure the surface of these terga and give them a fine dusty appearance. In this condition there is a distinct contrast between terga 3-4 and terga 1-2. At the other extreme, terga 3 and 4 have scattered, short, mostly simple hairs. Males exhibit variation in the number of hairs on all four anterior terga. The hairs are longer, more erect than in the female and do not vary noticeably in length and branching. Valley specimens of both sexes usually exhibit the condition with many hairs and the montane specimens the condition with few short hairs; individual variation partly obscures this geographic pattern. Specimens from the northern part of the range outside of California are intermediate, but tend to be closer to the valley condition. In southern California both valley and montane conditions exist as populational rather than geographic differences.
The female abdomen (in dorsal view) varies in width. Variation in abdominal width shows much the same type of geographic distribution as the variation in abdominal hair pattern mentioned above. Specimens from the valleys tend to have a wide abdomen, while montane specimens tend to have a narrower abdomen, In both sexes the integument usually gives off slight metallic reflections. In the female the color tends to be greenish-gold and in the male bluish-black. This is the common condition found, except in the mountains of California. The montane specimens are pure black without metallic reflections. In addition to this general geo¬graphic trend there is some individual variation. As a result either metallic or nonmetallic specimens can occur almost anywhere within the range of the species.
Both sexes vary in size, tending to be largest in the central valleys of California. In other parts of the range (both within and outside of California) specimens tend to be smaller in size, but at any one locality there may be a good deal of size variation.
The female hind tibiae appear to vary in width at the apex. This character is difficult to quantify and differences may be influenced by specimen size and curving of scopal hairs. Specimens from the mountains of California tend to have slightly narrower tibiae than is found in other parts of the range (both within and outside of California). Again individual variation has obscured any geographic trend which may occur.
The hairs along the anterior margin of the scopa are often directed outward. At other times the hairs curve posteriorly at their tips and overlap more posterior hairs. Thus scopal hairs tend to be orientated in the same direction and obscure the surface of the tibia. The second condition reaches its extreme in some southern California specimens, but specimens from other areas, especially in the mountains of California, may closely approach this condition. Individual variation is common in this character.
Males from most areas have the clypeus mostly yellow. Only the extreme basal margin and lateral areas are black. In the mountains of California the yellow maculation is very irregular in shape and much reduced in size, so that half or less of the clypeus is yellow. Individual variation masks all but a general geographic trend in which montane specimens show a reduced yellow maculation over valley specimens.
The male genitalia vary a great deal. Over most of the range the genitalia are near to figures 41-43. Notice the short, broad, dorsal lobes of the gonocoxite; the wide apex of the gonocoxite; the lack of large lobes on the inner margin of the gonocoxite; the large humped lamellae at the apex of the penis valve; the rather narrow expanded part of the penis valve; and the broad tip of the penis valve. In the mountains of California the genitalia look more as in figures 44-46. In spite of the marked differences in the genitalia many intermediates are found, especially in southern California. Here specimens have genitalia of an intermediate or mountain type. Different populations show all sorts of combinations of character states. Even populations in close geographic proximity may be quite different from each other.
In the characters discussed above the very few specimens from Utah, Nevada and Arizona would seem to fit the kind of variation shown in southern California.
The variation discussed thus far has been of the type which previously led to the splitting of chlorogaster into several species and subspecies. The reasons for not recognizing these forms is discussed for each.
Several characters seem to vary among individuals and do not show populational trends. The middle segments of the male antennae may vary in length among individuals, thus affecting the overall length of the antenna. The middle segments vary from slightly longer than broad to one-third longer than broad. The underside of the flagellum in both sexes varies in color; in the female the color is orange to dark brown and in the male yellow to brown. Some males exhibit varying amounts of black hair on the face and scape, while other males have all white hairs on these areas. The variation in this character may be partly due to age, the older specimens being lighter in color. In both sexes the pterostigma and wing veins exhibit various shades of amber, and the first transverse cubital vein may end in the pterostigma or end as much as two vein widths from the pterostigma. There may be just a few fine rugulae on the anterior part of the enclosure of the propodeum or the rugulae may extend posteriorly and cover most of the center of the enclosure.
|
|
|
Names | |
|
|
| Supported by | |
Updated: 2024-04-18 18:48:10 gmt
|