|
Reprinted with permission from: Daly, H. V. 1973. Bees of the genus Ceratina in America North of Mexico (Hymenoptera: Apoidea). University of California Publications in Entomology 74: 1-114. The Regents of the University of California. Published by the University of California Press.
Ceratina acantha Povancher, 1895
Female.Measurements and ratios: wing, 4.05 mm; hamuli, 5; eye, 95:110:110:110; clypeus, 50x95:30:85; interocellar, 22:42:20:50, Ø 15.0; frontal carina, 65; malar area 0x47; in teralveolar, 27:25:42:50, Ø 15; antenna, 45:12:12:7:7:7, Ø 13; subpleural signum, 15x2.5 (compared with lectotype; California, lion Angeles Co., Altadena; Univ. Kansas).
Structure: Head subquadrate in cephalic view; eyes parallel, narrowly rounded above, sub- truncate at malar area; frontal carina simple, defined only between alveolae, absent beneath median ocellus; vertex and supraorbital area limited posteriorly by preoccipital ridge without a carina; median portion of hypostomal carina when viewed laterally of low uniform height, gently terminating anteriorly without a tooth, when viewed ventrally angle between median and lateral portions about 120°. Basitibial plate spiniform, its elevated apex at basal 0.28 of tibial length. Metasoma widest at segments 34; gradulus of tergum 4 terminating laterally beyond spiracle at a distance of 1.3 spiracle lengths; tergum 6 with median central area flattened; graduli of sterna 23 slightly arcuate.
Sculpture and vestiture: Head shiny between punctures, largely impunctate clypeal disc, lower median supraclypeal area, lowest paraocular area, subantennal area, convex portion of alveolocellar area and in narrow band behind eye; sparsely punctured on mandible base and gena; elsewhere with fine to very fine punctures usually more than a puncture width apart and with largely simple setae; punctures closer in interalveolar, supraorbital and paraocular areas; sparse setae longer in interalveolar area, vertex, labral disc, postgena, and apically on mandible. Sides of pronotum finely punctured about a puncture width apart, impunctate area before pronotal pit; scutal disc shiny, virtually impunctate except sparsely along lines, fine punctures marginally and almost a puncture width apart laterally and on axilla; scutellar disc sparsely punctate, fine punctures becoming closer to about a puncture width apart laterally and posteriorly; metanotum finely punctured throughout, about a puncture width apart or more; mesopleuron with fine punctures more than a puncture width apart laterally, becoming more widely spaced posteroventrally, setae narrowly plumose, becoming longer posterolaterally (resembles fig. 84); hypoepimeral area impunctate in lower third; metapleuron finely punctured, about a puncture width apart or more; small impunctate area before metapleural pit; propodeal triangle reticulate, with longitudinal carinulae only mediad; angle of propodeum impunctate, minutely coriaceous; sides finely punctured, becoming closer and smaller laterad, with long erect, narrowly plumose setae. Metasomal tergum 1 finely punctured on dorsal surface more than a puncture width apart, virtually impunctate on anterior surface; terga 25 with pregradular area nearly impunctate on anterior terga, sparsely punctured on posterior terga; postgradular punctures fine, sparse on discs of more anterior terga, becoming closer to about two puncture widths apart laterally and apically, each with a short appressed simple seta; ground shiny, subapically minutely roughened only on tergum 4; tergum 5 sparsely and finely papillate; tergum 6 finely punctured about a puncture width apart, ground shiny between sparse low papillae which become rugulose laterally and apically; setae short, simple, except central area which is punctured but shiny and free of papillae (lateral view resembles fig. 11e). Sterna finely punctured, about two puncture widths apart or more; setae simple, fine, suberect, dense at apex of abdomen; setae of wax glands appressed.
Color: Body metallic blackish green (ISCC-NBS 152) with minor yellowish reflections; metal. lie colors lacking on labrum, mandibles, mesoscutal disc, and wax glands. Ivory marks: irregular amygdaliform on clypeus, extending 0.60 of clypeal length; spots at bases of all tibiae, being largest on metatibia. Pronotal lobes dark. Antennae fuscous, lighter beneath; legs proximally fuscous, femora and tibiae faintly metallic, tarsi apically testaceous; wing membrane testaceous, venation and tegula darker. Body setae pale flavescent.
Male.Measurements and ratios: wing, &7 mm; hamuli, 5; eye, 95:105:85:75; clypeus, 50 × 75:30:70; interocellar, 20:35:15:40, Ø 15.0; frontal carina, 50; malar area 0 × 30; interalveolar, 20:15:35:40, Ø 15; antenna, 32:12:11:7:10:15, Ø 15; subpleural signum, 15.0x2.5 (Mts. near Claremont, Los Angeles County, California).
Structure: Head rounded in cephalic view; eyes convergent below, narrowly rounded above, broadly rounded at malar area; frontal carina simple, distinct between alveolae, sulciform beneath median ocellus; vertex and supraorbital area limited posteriorly by preoccipital ridge with a low carina; median portion of hypostomal carina when viewed laterally of low uniform height, terminating anteriorly without a tooth, when viewed ventrally lateral portion arcuate. Scutum with all lines obscure; ventral profile of metatrochanter right-angled, apical anteroventral facet partly developed, minute punctures; width of metafemur at tooth 0.6 length, tooth apex narrowly rounded, acute 75° angle, basal facet absent, polished, ventral edge with tuft of medium setae at tooth apex (resembles fig. 10!). Metasoma widest at segment 4; gradulus of tergum 4 terminating laterally beyond and below spiracle at a distance of 2.0 spiracle lengths; tergum 6 without median subapical protuberance (resembles fig. lint); dorsal view of apical plate width 1.4 times length, narrowly rounded; sternum 6 with one median tooth partly divided at apex (resembles fig. 13g). Genitalia presumed to be similar to fig. 29g.
Sculpture and vestiture: Head shiny between punctures, largely impunctate on clypeal disc, lowest parocular area, subantennal area, most of convex portion of alveolocellar area and in narrow band behind eye; sparse fine punctures on mandible base; gena with fine punctures about a puncture width apart; elsewhere with fine to medium punctures about a puncture width apart and with largely simple setae; punctures closer in interalveolar area; sparse setae longer in interalveolar and postgenal areas, vertex, labral disc and posteriorly on mandible. Sides of pronotum finely punctured about a puncture width apart, impunctate area above pronotal pit extending to beneath dorsolateral angle; scutal disc shiny, virtually impunctate in center, but moderately punctured along lines, fine punctures marginally, laterally, and on axilla about a puncture width apart; scutellar disc impunctate in center, fine punctures becoming contiguous laterally and posteriorly; metanotum finely and contiguously punctured throughout; mesopleuron with fine punctures more than a puncture width apart laterally, becoming more widely spaced posteroventrally, long plumose setae; hypoepimeral area virtually impunctate in lower half; metapleuron finely punctured, about a puncture width apart; small impunctate area before metapleural pit; propodeal triangle subreticulate rugulose, longitudinal carinulae distinct even laterally; angle of propodeum impunctate, minutely coriaceous; sides finely punctured, becoming contiguous and smaller laterad, with erect, plumose setae. Metasomal tergum 1 finely punctured on dorsal surface about two puncture widths apart, virtually impunctate on anterior surface; terga 26 with scattered fine punctures on pregradular area near gradulus, postgradular punctures fine and uniformly about two or more puncture widths apart, each with a short appressed simple seta; ground shiny, but minutely roughened on terga 46; tergum 7 finely punctured about a puncture width apart with shiny ground, apical plate above with long, spreading plumose hairs extending beyond plate. Sterna 15 finely punctured, more than a puncture width apart; setac fine, suberect, narrowly plumose, longer subapically on each sternum.
Color: Head and thorax metallic very dark bluish green (ISCC-NBS 166); abdomen similar but cuticle beneath metallic reflections dark fusco-rufous; metallic colors lacking on mandibles which are dark fusco-rufous. Ivory marks: broad hat-shaped on clypeus, extending full length and width; subrectangular spot on labral disc; spots at bases of all tibiae, being largest on metatibia. Pronotal lobes dark. Antennae fuscous, lighter beneath; legs proximally almost black, femora and tibiae faintly metallic, tarsi apically fuscous to dark testaceous; wing membrane hyaline, venation and tegula dark testaceous. Body setae nearly white.
Male compared with illustrations of other specimens: metafemur and metatrochanter (fig. 10!), lateral view of segments 67 (fig. lint), sternum 6 and apical plate (fig. 13g); genitalia not dissected but illustrated from another specimen (fig. 29g). The lectotype designated here for C. acantha is a female specimen from the Provancher collection bearing a top yellow label with the number 1711, a second label with a double red margin Ceratina acantha Prov., Cal., a third red label Lectotype 247, Ceratina acantha Provancher, Comeau, 1944, and my red lectotype label. Noel M. Comeau apparently never published his designation. The specimen is deposited in the Provancher collection, Université Laval, Quebec, Canada. It has a reduced clypeal mark and wing length 3.9 mm. The lectotype designated here for C. submcsritim4 is a male specimen bearing a top label 5-13-96, a middle label Seattle, Wash., Lot 214, and will be deposited in the Academy of Natural Sciences of Philadelphia. The specimen apparently was among those examined by Cockerell because he states I have also before me a couple from Seattle, Wash., May 13th, from Mr. Dunning, marked Lot 214.
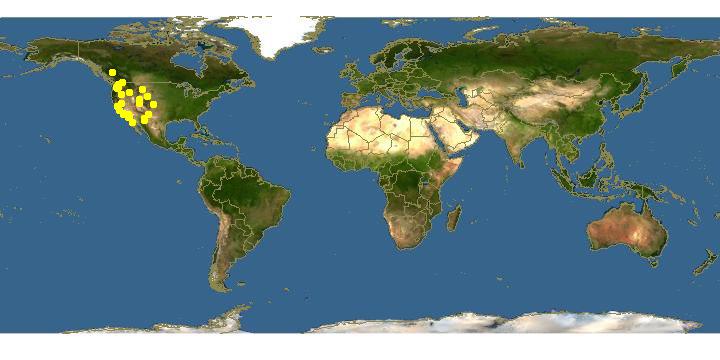
Ceratina acantha is the most commonly collected species of small carpenter bee in the western United States (fig. 14). The unusually dark, shiny, metallic green or blue-green color; dark pronotal lobes; fine, sparse mesopleural punctation; and medium size will identify most females. The male is similarly characterized and also has the unique large, acute, ventral tooth on the hind femur with the apex of the tooth closer to the apex of the femur than to the base. Small females of C. tejonensis or C. micheneri resemble C. acantha, but may be distinguished by their coarse mesopleural punctation. Some C. acantha have varying amounts of ivory color at the apex of the pronotal lobe and may be easily confused with C. nanula. The mesopleural punctures of the latter are larger and only a puncture width apart or less. The sixth metasomal tergum may also be examined; it is roughened and dull in C. nanula and usually glistening in C. acantha. Small males of C. micheneri may be separated from C. at ant ha by their possession of a shiny basal facet on the ventral tooth of the hind femur, a single median tooth on the sixth sternum, and the coarse mesopleural punctation.
This species is sufficiently abundant that preliminary analysis of both phenetic and biological variation is possible. It is exceptional biologically because several populations in Los Angeles County reproduce by thelytokous parthenogenesis. This mode of reproduction is rare among bees, being known in certain races of Apis mellifera Linnaeus (Tucker, 1958) and in the alien Ceratina dallatorreana Friese (Daly, 1966b). The near absence of males of C. acantha in Pasadena, California, was first noted by Michener (1936).
Sex ratios vary widely in collections from throughout the distribution. Male specimens may be lacking from poorly collected areas because they naturally decline in frequency during late spring and early summer. For example, bisexual populations in nests at Davis, California, range from 28.7% males in November, 11.4% males in late April, to 2.9% males in mid-June (Daly, Bohart, and Thorp, 1971). The proportion of males computed for larger collections from various localities are: Berkeley and Oakland (176 ♀ ♀, 148 ♂ ♂, or 45.7% ♂ ♂), Contra Costa County (70 ♀ ♀, 75 ♂ ♂, or 51.7% ♂ ♂), Los Angeles County (207 ♀ ♀,14 ♂ ♂, or 6.3% ♂ ♂), Riverside (64 ♀ ♀, 29 ♂ ♂, or 31.2% ♂ ♂, Davis (80 ♀ ♀, 80 ♂ ♂ , 50% ♂ ♂), Corvallis (77 ♀ ♀, 63 ♂ ♂, or 45% ♂ ♂), and Seattle (87 ♀ ♀, 66 ♂ ♂, or 43.l% ♂ ♂).
The exceptionally low frequency of males from Los Angeles County was investigated by examining nest collections and sex ratios from restricted areas with adequate collections. Positive evidence of thelytoky was accepted if an unfertilized female with signs of parenthood was associated in a nest with all female offspring. The immatures were reared in the laboratory so that fortuitous associations and escape of males was avoided. Details of the general approach are described by Daly (1966b). Evidence of this kind was obtained in 1969 for C. acantha at the following localities: Santa Monica Mtns., 1 ml. s. Ventura-Los Angeles Co. line on State 23; Eagle Rock, north end of Argus Dr. and Hillside, 7601080 ft. elev.; Eagle Rock, intersection of Arbor Dell P1. and Wameda Ave. and hillside 840960 ft. elev.; Altadena, intersection of Loma Alta Dr. and Lake Dr., Los Flores Canyon, 18011920 ft. elev.; Glendale, intersection of Verdugo Blvd. and Calle Vaquero,720 ft. elev.; and Montrose, intersection of Verdugo Blvd. and Canon Rd., 1200 1220 ft. elev. in each case the entire sample from the population appeared to be thelytokous.
Circumstantial evidence was obtained that another thelytokous population existed in the Puente Hills. During 19201929 and at various seasons, P. H. Timberlake (in litt.) collected 50 females and no males from an area along La Canada Verde Creek about one mile in diameter and about two miles east and somewhat south of downtown Whittier. One male was collected at the river in Whittier. The females are virtually monomorphic in color, each with a small clypeal maculation. A sample of 45 wing lengths from this locality has the lowest variance of all populations measured (see below). No recent collections have been attempted.
Bisexual populations definitely exist in the coastal Palos Verdes Hills (fertilized females with offspring of both sexes collected in 1966 and 1969 from Marineland and near the intersection of Palos Verdes Drive and 25th St.), Santa Cruz Island (157 ♀ ♀, 161 ♂ ♂collected from nests 1417 March 1969), and probably at Riverside based on the museum collections. Elsewhere in the vicinity of Los Angeles, males have been occasionally collected, but the total numbers from any one location are insufficient to judge their relative abundance. At the present, all populations outside of Los Angeles County are assumed to be bisexual.
Phenetic variation has been previously recognized in C. acantha by the description of two additional taxa: C. submaritima and C. s. ehrhorni. The latter is based on a male which was said to be yellowish-green from San Jose, California. The type is no longer yellowish, but occasional specimens at various distant localities have faint brassy or yellowish reflections, in the absence of distinctive populations having this color, this rare variant is considered part of the normal variation of C. acantha. Michener (1936) divided the species on a population basis into a northern melanic subspecies, C. a. submaritima, and a southern subspecies, C. a. acantha, with more extensive clypeal maculations. This classification was re-examined by comparing two characters among twelve localities with collections sufficient to permit statistical analysis. Whig length for both sexes was chosen as a continuous character because it is highly correlated with general body size and the diverse allometric features which influence head shape. The measurements were analyzed by Powers modification (1970) of Gabriels (1964; Gabriel and Sokal, 1969) Sums of Squares Simultaneous Test Procedure. The measurements were also transformed to natural logarithms and sample variances recomputed to compare variability (Lewontin, 1966). Presence or absence of a clypeal maculation, no matter how small, was chosen as a meristic character because it provides a measure of the degree of melanization over the entire body which in turn affects the general body color. Analysis was made by a R x C test of independence using the G-test (Sokal and Rholf, 1969).
When the results of the separate analyses are examined certain broad trends can be perceived (tables 2 and 3). However, the complex phenetic relationships do not permit conventional taxonomic subdivisions to be recognized. In general the bees from the northwestern localities (Seattle and Corvallis) are members of the same character subsets and distinguished by their larger size and virtual absence of clypeal maculations. To the east the bees from distant localities (Arrow Junction and Logan) are likewise closely similar to each other and lack clypeal maculations, but are strikingly smaller. Average wing length also decreases from southern Oregon to southern California and the frequency of clypeal maculations increases. The phenetic relationships of samples in this region are complex, especially in southern California. No two samples are members of the same combination of character subsets and membership in a character subset is often unrelated to geographic proximity.
Two of the above samples were from populations thought to be thelytokous (Pasadena and Puente Hills). Phenetically they are most closely related to bisexual populations at Riverside and San Diego but they curiously rank in the analyses as the samples with the smallest wing lengths and highest frequency of clypeal maculations. However, these are not phenetic criteria by which thelytokous populations can be recognized. Inspection of the thelytokous bees from the Santa Monica Mountains shows that only 20% have clypeal maculations and average wing length ranks slightly larger (4.03, n = 9). Presumably each parthenogenetic population has been derived independently from a nearby bisexual population and largely preserves the original phenetics of the ancestor.
Comparison of the variances of the transformed wing lengths reveals an unusual relationship. At each locality the samples of males consistently rank less than the females and they are significantly less variable at four localities: Seattle (F = 2.62***), Klamath Falls (F 1.69*), Berkeley (F = 1.97*), and Santa Cruz Island (F = l.73**). This is surprising in view of the deductions by Eickwort (1969) that males should be more variable than females in arrhenotokous populations.
Ceratina acantha is collected in a great variety of vegetation types in the western United States (fig. Sc). Nests are most commonly found in woody shrubs or trees such as Sam bucus sp. (fig. 31a) or Ailanthus sp. (Daly, Bohart, and Thorp, 1971). The egg is placed at the rear of the pollen loaf (fig. 31o).
|