| ||||||
| ||||||
|
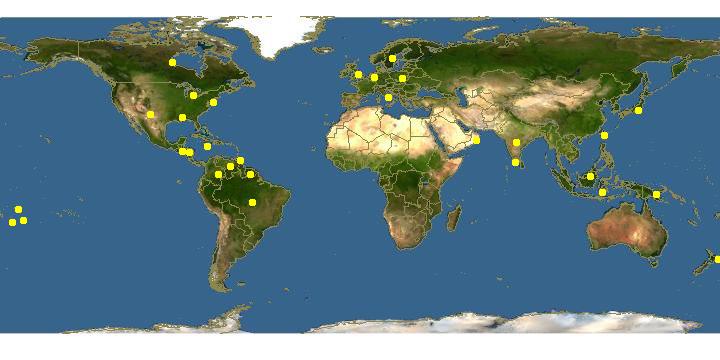
Ceratocystis fimbriata Ellis and Halsted MANGO BLIGHT Mango wilt; Cacoa wilt; Rubber mouldy rot; Sweet potato black rot; Coffee canker; Aspen black canker; Aspen target canker; Taro black rot; Plane tree canker stain; Mallet canker; Black Rot |

Updated: 2024-04-25 06:37:08 gmt
|