|
Reprinted with Permission from: R.J. Oram. 2018. Revision of the Genus Hylaeus Fabricius (Hymenoptera: Colletidae) in Canada. Masterâs Thesis University of Regina
Diagnosis: Females of Hylaeus leptocephalus are similar to females of H. communis. They can be distinguished from H. communis by the, the large irregular spaced punctures, with shiny interspaces in H. wootoni and large uniformly spaced episternal punctures, with tesselate interspaces in H. communis. Males of Hylaeus leptocephalus can be easily separated from other species by pale white maculations on the face, with the ocular maculations truncate on top, and the scape having white maculations on the outer edge.
Females: average face width = 1.48mm; average thorax length = 1.71mm; average intertegular width = 1.06mm; average forewing length = 3.33mm. (n=5) Black, except for white maculations on lower paraocular which extend from above the malar area to below the level of antennal sockets, collar sometimes with small lateral maculations (sometimes absent), apical half of pronotal lobes, hind tibia on basal 1/3,
tegula sometimes with a pale maculation on basal end, costal sclerite maculated; wing membranes clear, veins brown
Head: Scape straight, with dense, shallow punctures covering the surface, F1 quadrate, F2â3 broader than long, about half the length of F1, F4âF9 quadrate, subequal in length to F1, F10 slightly longer than broad; clypeus as long as broad, with shallow punctures (i
> 3pd), interspaces with deep grooves running vertically from the basal to apical edges; superclypeal area with shallow punctures dense (i < 1pd ) basally between antennal sockets, becoming sparser apically (i > 1pd); upper paraocular area and frons with coarse punctures (interspaces hardly visible); vertexal area weakly elevated, punctures with (interspaces hardly visible), with punctures becoming sparse between lateral ocellus and compound eyes (i < 1pd), interspaces smooth; compound eyes with inner margin weakly converging below; facial fovea short, with lower margin at level of bend of inner eye
margin, extending to upper curve of eye, fovea separated from the compound eye by the width of one fovea; face with sparse, short pale hairs, with long pale hairs at the top of the vertexal area; mandibles with long pale erect hairs.
Thorax: Pronotal collar rounded anteriorly, width of collar even throughout; mesoscutum with coarse and dense punctures (i < 1 pd), interspaces tesselate; parapsidal lines deep, about the length of the tegula; scutellum with deep punctures larger than mesoscutumn, (i
= 1pd), with interspaces tesselate; metanotum rugosopunctate;
mesepisternum with anterior and lateral faces rounded, punctures deep and scattered, interspaces less than or equal to 1pd with interspaces smooth; metepisternum rugosopunctate; propodeum with basal area heavily rugose, width subequal to metanotum, separated from posterior surface by a strong carina, posterior and lateral surfaces rugosopunctate, posterior surface separated from lateral surface by a weak carina; fore coxae rounded. Mesoscutum and scutellum with sparse short, erect, white simple hairs; metanotum with short, white hairs
medially, with elongate, white branched hairs laterally; pleuron with short erect white hairs, intermixed with sparse short erect dark hairs subequal to the hairs on the mesoscutumn; propodeum with short, white branched hairs on the posterior and lateral surfaces, with longer, branched hairs under the wing on posterior surface; foreleg with a
mixture of short and long white branched hairs on coxa, mixture of short erect pale simple hairs and long branched white hairs on femur, flattened pale hairs on tibia, long, dense pale hairs hooked apically on tarsi; mid leg with short branched white hairs on coxa, short pale hairs on femur, short, flat pale hairs on tibia, long, sparse pale hairs on tarsi; hind leg with short, semi-erect white hairs on coxa; with short, pale hairs throughout on remaining segments, with erect hairs intermixed on basitarsus.
Abdomen: T1 surface smooth and shiny, with small, widely spaced (i > 3pd) punctures, T2 âT5 smooth and polished, T1âT6 with small pits
from which short, fine pale white hairs arise becoming more dense laterally, T6 smooth and polished, with long, erect, dark hairs arising from apical edge; S1âS5 smooth, with small pits from which intermixed, short, pale, and dark hairs arise, S6 smooth; S5 with a line of sparse black hairs delimiting an apical golden crescent; S6 with long dark hairs.
Males: average face width = 1.67mm; average thorax length = 1.26mm; average intertegular width = 1.21mm; average forewing length = 3.43mm. (n=5)Black, except for white maculations on the clypeus, supraclypeal area, sides of face with maculation covering lower paraocular area extending from malar area to above the antennal sockets, maculation truncate at top, slightly separate from the eye, scape maculated on the outer edge, maculations on the pronotal lobes (well-separated medially), fore tibia maculated in full dorsally, fore basitarsus white (apical edge black), mid tibia on basal 1/3, basitarsus white, hind tibia with maculation on basal half,
basitarsus white (apical edge black), tegula with a small maculation basally, costal sclerite maculated; wing membranes slightly tinted brown and clear, veins brown.
Head: Scape width subequal to the width of the flagellum, with dense, shallow adpressed punctures covering surface, F1âF2 slightly broader than long, F3âF10 quadrate, F11 slightly longer than broad; clypeus with length equal to width with sparse, extremely shallow punctures (i = 3pd) and interspaces smooth; supraclypeal area long, length subequal to clypeus, impunctate; upper paraocular area and frons with punctures coarse and dense (interspaces hardly visible); vertexal area slightly elevated, densely punctured (interspaces hardly visible); compound eyes with inner margin weakly converging below;
facial fovea extremely short, localized at the upper edge of compound eye not diverging; face with short, erect, pale hairs; mandibles with short erect hairs; genal area with variable pale hairs.
Thorax: Pronotal collar rounded anteriorly, width of collar even throughout; mesoscutum with coarse and dense punctures (i < 1 pd), interspaces tesselate; parapsidal lines deep, about the length of the tegula; scutellum with deep punctures larger than mesoscutumn, (i
= 1pd), with interspaces tesselate; metanotum rugosopunctate; mesepisternum with anterior and lateral faces rounded, punctures coarse (i = 1pd), interspaces weakly tesselate; metepisternum rugose; propodeum with basal area heavily rugose, width equal to metasuctum, separated from posterior surface by a strong carina, posterior and lateral surfaces, with shallow close punctures, with interspaces hardly visible, posterior surface separated from lateral surface by a weak carina; fore coxa rounded; mesoscutum with short, erect, pale simple hairs; scutellum with long, erect, pale simple hairs; metanotum
with long, pale white hairs medially, with long, white, branched hairs laterally; pleuron with long, white, simple hairs; propodeum with short, white branched hairs at edge of posterior surface, with long, white branched hairs under the wing base on posterior and
lateral surfaces; foreleg with long white branched hairs on coxa, short pale erect hairs dorsally and long pale erect hairs ventrally on femur, flattened pale hairs on tibia, long, dense pale hairs on tarsus and basitarsus; mid leg with short simple hairs throughout, with
pale erect hairs on the tarsus and basitarsus; hind leg with short, semi-erect branched, white hairs on coxa, with short, pale hairs throughout on remaining segments, with erect hairs intermixed on basitarsus.
Abdomen: T1 with surface smooth and shiny, with small, widely spaced (i > 3pd) punctures laterally, T2 âT7 with larger punctures than T1 (i = 2pd) with interspaces smooth, T1âT7 with small pits from which short, fine pale white hairs arise causing the surface to look roughened, T6 with short, erect white hairs on apical edge, T7 with short erect white hairs; S1âS6 smooth, with small pits from which small fine pale white hairs arise, S5 with short erect white hair on apical portion, S6 with short, erect, white hairs.
Reprinted from: Snelling, R. 1970. STUDIES ON NORTH AMERICAN BEES OF THE GENUS
HYLAEUS. 5. THE SUBGENERA HYLAEUS. S. STR. AND
PARAPROSOPIS (HYMENOPTERA: COLLETIDAE) Contributions in Science, No. 180.
This species is not closely related to any other in the Nearctic fauna, but
is very similar to the Palearctic H. bisinuatus Forster. The two are so closely
related that I know of no reliable external feautres which will separate them.
The male terminalia of the two are very similar to one another. More material
must be studied, however, to determine whether they are conspecific or not.
This species has recently been introduced into Hawaii (Beardsley, 1959),
and is evidently adventive in parts of its range where it is now quite common.
I believe that this species has extended its range appreciably within the past
50 years. The earliest known capture of this species is in 1912 at Fargo, North
Dakota; these specimens form the type series. The species has been collected
in North Dakota on many occasions since then, but remained rare in Hylaeus
samples taken until after 1925. After this date it became increasingly common,
until it is now the species most frequently captured.
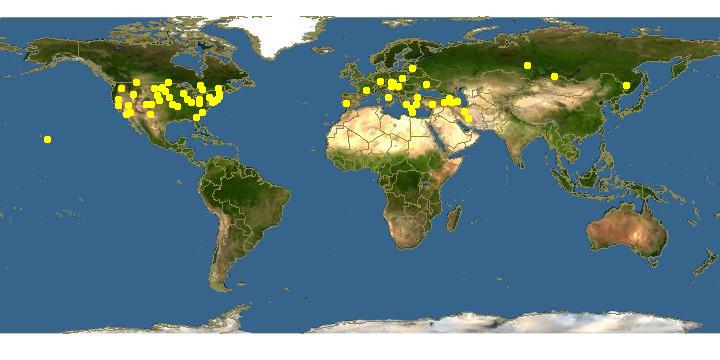
The earliest records outside of North Dakota are 1915 (Colorado) and
1916 (Alberta, Canada). Earliest records for other states, proceeding west
from North Dakota to the Pacific coast, are: Wyoming and Idaho (1920),
Utah (1922), British Columbia and Washington (1925) and California
(1933). This species is found in eleven states east of the Mississippi River; the
earliest date of capture which I have seen is 1927 in Massachusetts. In none
of his numerous papers on eastern bees does Robertson mention this bee; this
species is so distinctly different from the remaining eastern Hylaeus that he
certainly would have recognized it as different from any previously described.
Within California the earliest record for this species is 1933 at Sacramento.
There are few records in the 1930's and 1940's. But, beginning with
1950 there are abundant records, largely from the Central Valley area. My
greatest field acquaintance with this species is in California, so the following
observation must be considered with this point in mind. All of the California
records are within, or on the periphery of, agricultural cultivated areas. I can
affirm, further, that the bulk of my records from most of the area west of the
Continental Divide come from similar situations, as nearly as I have been able
to determine.
This bee is of further interest in that it alone of all the species of Hylaeus,
s. str., exhibits a pronounced tendency toward oligolecty on flowers of the
family Leguminosae. Of 87 specimens bearing floral data, 56 were taken on
Melilotus species. These legumes are introduced from the Palearctic region.
Of the remaining 31 records, 13 are from introduced Palearctic plants in other
families (Amaranthaceae, Umbelliferae, Cruciferaceae, Tamaricaceae, Leguminosae) and eight of these are records of males. The remaining 18 records
involve the families Polygonaceae, Asclepiadaccae, Salicaceae and Compositae,
all belonging to genera represented in the Palearctic region: six of these
records are of males.
The above data would seem to indicate the possibility that this species was
introduced into the area around Fargo, North Dakota sometime prior to 1912.
Once established there it began to extend its distribution both east and west
and by 1925 apparently reached both coasts; subsequent distribution has been
extended southward, so that this species now occurs in southern California,
Oklahoma and Georgia. It is now a well-established part of our fauna and
appears to be oligolectic on the legume genus M eli/ollis. If this is, in fact, an
introduced species, it may well prove to be a synonym of a described Palearctic
species. This, however, may not be decided for many years yet due to the
extremely chaotic status of the taxonomy of European Hylaeus.
It is possible that this species is a native component which was, prior to
the first quarter of the century, a very rare species. Subsequent to that time the
increased abundance of a suitable food-source plant (the introduced genus
Melilotus) and the conditions of an unbalanced ecology may have permitted
this species first to thrive in its native region and then to spread out to other,
similarly disturbed areas. Of numerous series of specimens reared from soda
straw traps by A. R. Gittins in Idaho, many are referrable to H. stevensi. This
species, then, is strongly apt to exploit artificial nesting possibilities, such as
exist in and around farm buildings.
|
Reprinted with permission from: Mitchell, T.B. 1960 Bees of the Eastern United States. North Carolina Agricultural Experiment Station Technical Bulletin No. 141.
FEMALE�Length 5-6 mm.; black, antennae piceous above, more testaceous beneath; face marks pale yellow, filling area between clypeus and eyes, abruptly truncate at level of antennae; tubercles, transverse marks on collar, tegulae anteriorly, pale yellow; posteriorly the tegulae are ferruginous; wings very lightly infuscated, veins and stigma brownish; tibiae yellow at base, legs otherwise dark; face narrowed below; cheeks slightly more than half as broad as eyes in lateral view; facial foveae rather short and obscure, linear, separated from eyes by about an equal space, not at all divergent above; basal segment of flagellum about as long as broad, 2nd and 3rd segments distinctly broader than long, 4th nearly as long as broad, the following segments fully so; punctures of face below antennae shallow, obscure and sparse, more coarse, deep and distinct above antennae, well separated but not sparse; very coarse and deep on thorax above, quite close over most of scutum, somewhat more sparse on scutellum, and even more coarse and close on pleura; dorsal area of propodeum quite extensive, hardly distinguishable from metanotum, both very coarsely rugose; abdomen shining, very finely but distinctly punctate, rather sparsely so on basal segment, closer on the more apical segments.
MALE�Length 4-6 mm.; black; antennae pale ferruginous, scape blackish, maculated; tegulae more piceous, maculated; wings lightly infuscated, veins and stigma brownish-piceous; mandibles and labrum black; maculations cream-colored, as follows: entire face below antennae, supraclypeal mark extended upward between antennae, lateral face marks obliquely truncate above antennae, very slightly divergent from eyes above, narrow anterior stripe on scape, large blotch - on tegulae, transverse marks on collar, tubercles, front tibiae in large part, and apex of femur, basal portions of mid and hind tibiae, and mid and hind basitarsi; other tarsal segments, including entire front tarsi and anterior face of front tibiae more ferruginous; face narrow, elongate; eyes convergent below; cheeks about half as wide as eyes in lateral view; scape slender, no broader than flagellum, slightly curved, basal segment of flagellum considerably broader than long, length and breadth of 2nd and following segments subequal; front coxae simple; dorsal area of propodeum rather extensive, hardly distinguishable from metanotum, both being very coarsely rugose, scutum and scutellum shining, very coarsely and deeply punctate, punctures well separated but not sparse, punctures on pleura even more coarse and quite close; abdomen deeply and distinctly, although relatively finely punctate, rather sparsely so on basal segment, closer on the more apical segments.
DISTRIBUTION�In the East this species has been collected in Michigan, New York, New Jersey, Indiana, District of Columbia, Virginia, North Carolina and Georgia, as also in Quebec and Ontario. It is in flight from May until September.
FLOWER RECORDS�Only one flower collection record has been made, one specimen having been collected on cow-pea (Vigna inensis), at Raleigh, North Carolina.
|