|
80x5 -
240x3 -
240x4 -
320x1 -
320x2 -
320x3 -
640x1 -
640x2
Set display option above.
Click on
images to enlarge. |

© Copyright source/photographer
· 9
Bombus sandersoni, Mid-Atlantic Phenology |

© Copyright source/photographer
· 7
Bombus sandersoni, f, back, Giles Co, VA |
|

© Copyright source/photographer
· 7
Bombus sandersoni, f, face, Giles Co, VA |

© Copyright source/photographer
· 7
Bombus sandersoni, female, black hairs posterior scutum |
|

© Copyright source/photographer
· 7
Bombus sandersoni, female, face |

© Copyright source/photographer
· 7
Bombus sandersoni, female, side |
|

© Copyright Laurence Packer 2014
· 7
Bombus sandersoni FEM comp |

© Copyright Laurence Packer 2014
· 7
Bombus sandersoni MALE comp |
|

© Copyright source/photographer
· 5
Bombus sandersoni, female, back |

© Copyright source/photographer
· 5
Bombus sandersoni, female, behind3 |
|

Nancy Lee Adamson · 1
Bombus sandersoni, female |

Nancy Lee Adamson · 1
Bombus sandersoni, female2 |
|

Nancy Lee Adamson · 1
Bombus sandersoni, female3 |

Nancy Lee Adamson · 1
Bombus sandersoni, female4 |
|
Overview |
Reprinted with permission from: Mitchell, T.B. 1962 Bees of the Eastern United States. North Carolina Agricultural Experiment Station Technical Bulletin No. 152.
QUEENLength 15-16 mm., breadth of abdomen 6 mm.; black, the more apical segments of the legs becoming brownish-testaceous, mid and hind spurs testaceous, tegulae brownish-piceous; wings lightly infuscated, being almost hyaline in central area, veins pale testaceous to piceous; pubescence copious but rather short, largely pale on head, with long, erect, black hairs on vertex and front, a small amount of short, greyish, densely plumose hairs around antennae, cheeks below becoming somewhat more greyish pubescent, and a few pale hairs intermixed with the dark on vertex; pubescence dense, erect and yellow on pronotum, tubercles, narrow anterior margin of scutum, most of pleura, propodeum laterally, and entire scutellum, that on venter somewhat more greyish, largely fuscous on legs but largely pale on coxae and trochanters, mid trochanters with a patch of short, curved, brownish bristles beneath; front and mid tibiae with rather elongate pubescence posteriorly and on outer face, the corbicular fringe on hind legs of very long, dark hairs; tarsal segments thinly covered with very fine, pruinose hairs that do not obscure the surface, basitarsi beneath with brownish pubescence; abdominal terga 1 and 2, and sometimes 5 and 6, yellow pubescent, with no dark mixture on 1 or 2, terga 3 with erect and copious black pubescence; clypeus quite finely, closely and deeply punctate laterally and above, broad median area shining and nearly impunctate; mandible with a basal ridge which is somewhat interrupted medially, this area somewhat depressed, its apex very slightly elevated above margin on each side, and fringed with short, ochraceous hairs; apex of mandibles with a pair of distinct and rather deep notches toward upper angle, very slightly and rather broadly emarginate toward lower angle, outer face toward base smooth and somewhat shining, with only minute and very vague punctures; malar space shining and impunctate, slightly shorter than basal width of mandible, about one-fourth length of eye; punctures over median area of face very fine and rather densely crowded, becoming more distinct but still quite close toward ocelli, largely shining and impunctate between ocelli and eyes; vertex medially with very fine, densely crowded punctures, these becoming somewhat more distinct but still very fine and close laterally; lateral ocelli subequally distant from eyes and margin of vertex, somewhat more widely separated from each other; antennal scape somewhat more than half the total length of flagellum, basal segment of flagellum considerably longer than either 2 or 3, but somewhat shorter than the two combined; hind basitarsus almost parallel-sided, posterior margin only very slightly curved, median length nearly twice the breadth; tergum 6 somewhat shining, very minutely and rather sparsely punctate medially, more closely so toward margins, apex narrowly rounded, very densely short pubescent, forming an apical tuft.
WORKERLength 8-13 mm., breadth of abdomen 3.5-5.5 mm.; resembles queen, but pubescence relatively more elongate, with the same general pattern except that the apical abdominal terga are usually entirely black pubescent.
MALELength 10-13 mm., breadth of abdomen 5-6 mm.; pubescence long and copious, in large part yellow on head, especially on face below antennae, on vertex medially and cheeks posteriorly, with intermixed, long, black hairs along inner orbits, around and above antennae, behind ocelli, on cheeks above and along posterior orbits; center of scutum thinly pubescent, usually posterior half black, the scutellum, pleura and anterior half of scutum yellow; legs largely yellow pubescent basally, hairs long and quite copious, tibiae with somewhat shorter pubescence on outer face, fringed with pale hairs, more or less intermixed with black, the mid and hind basitarsi with brownish-ochraceous pubescence beneath, largely bare on exterior surface, fringed with ochraceous hairs; hind tibiae quite slender, outer surface shining but convex, with sparse punctures, fringed with dark hairs anteriorly, and with more elongate, pale hairs, intermixed with dark, posteriorly; abdominal terga 1 and 2 with copious and elongate yellow pubescence, following terga black; clypeus with fine and close but quite distinct punctures beneath dense yellow pubescence; labrum rather broadly truncate, smooth and shining, with only minute scattered and rather obscure punctures; mandibles slender, bidentate apically. densely covered with ochraceous pubescence on outer face, this forming an elongate, somewhat curled fringe beneath; malar space smooth and shining, with only very vague and obscure punctures, length somewhat greater than basal width of mandible, about one-fourth length of eye; median area of face beneath the dense pubescence very finely and closely punctate, the punctures becoming distinct and quite sparse above toward ocelli, space between ocelli and eyes shining and impunctate, vertex medially quite closely punctate becoming more sparse and irregular laterally; lateral ocelli subequally distant from eyes, margin of vertex and each other; basal segment of flagellum very slightly longer than segment 2, about equal to segment 3; median width of hind basitarsus slightly less than one-third the length; sterna 7 and 8 and genital armature similar to impatiens (fig. 133).
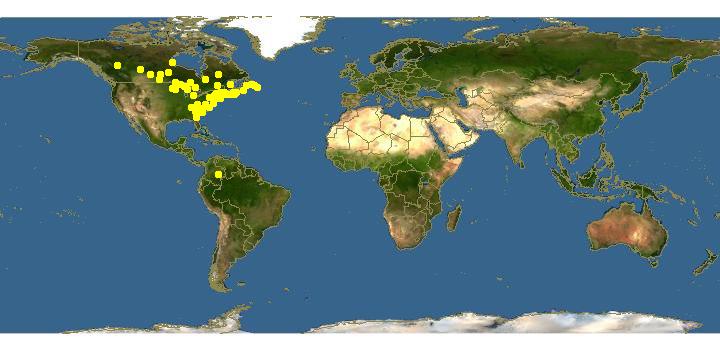
DISTRIBUTION Ontario to Newfoundland, south to Tennessee and North Carolina, April to October.
FLOWER RECORDSEpilobiurn, Mat- us, Pentsternon, Rhododendron, and Rubus.
Although sandersoni was originally described as a form of vagans, it has more recently been placed under frigidus (Catalog of Hymenoptera, p. 1252). However, Milliron has expressed the opinion in correspondence that it is distinct and that neither typical frigidus nor any of its forms occurs in the East.
|
|
|
Kinds |
Extracted from A MORPHOMETRIC STUDY Of' THE TAXONOMlC STATUS
OF BOMBUS SANDERSONI (HYMENOPTERA: APIDAE) by Plowright C., R. (1978).
Colour Patient S.
The 21 Jrigidus specimens show little chromatic variation In all, the thorax is \
yellow with a well-marked black intenilar band, abdominal terga I and 2 bear yellow \
pile, tcrgum 3 is uniformly black and terga 4-6 are clothed with orange or
yellowish-orange hairs. Specimen No. 14 (Table 0 differs from the rest in possessing a J
markedly yellow face but individuals with Ihis character occur fairly commonly in /
collections o(frigidus from all parts of its range ^/
The colour pattern of sandersoni, on the other hand, is extremely variable. Among the 36 specimens examined in this study, the colouration ranges from that typical of vagans (the Franklin cotypes. Nos 14 & \5) to individuals which are chromatically indistinguishable from jrigidus except for the absence of yellow or orange pile on T4. Even this latter character breaks down partly in the case of the Cresson couperi lectotype (No. 36), which has T4 black except for a posterior fringe of orange hairs. The chromatic variability in sandersoni may be appreciated by inspection of Table III, though we should also mention that we have before us many other specimens from
Ontario and New Brunswick which show all possible intergrades between ihe colour configurations represented in the group of 36 measured individuals,
Length of Malar Space andAntenna} Segments 3-5
Lengths of malar spaces in B. vagans ranged from 0.SOU) 0.86 mm (mean = 0.832
mm) and do not overlap with either frigidus (range: 0.50-0.60 mm, mean - 0.525 mm)
or sandersoni (range. 0.41-0.66 mm, mean = 0 535 mm). Mean malar space length of I frigidus is not significantly (P > 0.05) different from that of sandersoni. The greater malar length of vagans has, as mentioned previously, been the primary structural character used to separate this taxon from sandersoni. Most individuals can readily be placed in one or the other group simply by visual inspection of the shape of the face: in vagans the face is noticeably long in comparison with the square appearance of the faces of sandersoni and frigulus.
However, part of the difference between the malar lengths of vagans am sandersoni is accounted for by the generally larger body size of the former species (P < 0.01 for our samples). We therefore expressed the malar length data in ratio to (II the length of the 3rd antcnnal segment, (2) the length between the intersections of (a) the transverse median and anal veins and (b) the 3rd intercubital and marginal veins (the distance OR-S in Plow rig lit and Stephen, 1973). Both ratios effectively separate vagans, on Ihe one hand, from frigulus and from sandersoni on Ihe other (Table IV), but neither serves to discriminate between jrigidus and sandersoni.
Wing Venation
The scores of all specimens on the two discriminant axes are plotted in Fig. 1. Good separation is achieved between each of the 3 taxa. Both discriminant functions arc highly significant (P < 0.0001).
Principal components analysis yielded an imperfect separation between sandersoni |
and frigidus (Fig. 2), The first two roots of the correlation matrix accounted for 25.6% J
and 14.7% of the trace. Inspection of the positions of the specimens on the first two I
principal axes reveals no association between body size or geographic origin with either f
of these two components
The Cresson lectotype of B. couperi (No. 36) falls in an ambiguous position. On the principal components plot this specimen is situated in the overlap zone between frigidus and sandersoni, and although the discriminant analysis places it clearly within the sandersoni group (Fig. 1). it Iscs nearer to the frigidus cluster than any of the sandersoni specimens The two Franklin sandersoni cotypes (Nos. 14 & 15), by pontrast, appear to be more representative of the sandersoni group as a whole.
Discussion
Although our findings provide further evidence in support of Frison's (1929) intention that couperi ( = sandersoni) is structurally distinct from vagans, the relationship between sandersoni and frigidus remains somewhat obscure.
The case for separating sandersoni and frigidus seems, at first sight, to be sustained by the discriminant analysis results (Fig. 1). The fact that principal ■components analysis fails to separate the two forms into non-overlapping clusters can be
^amw^wwjwwjhw*
652
T11E CANADIAN ENTOMOLOGIST
explained away by invoking the argument advanced by Robinson and Hoffmann {I975): there is no reason to expect that any of the principal axes should be closely aligned wjih those axes which express differences due to recent phylogenetic divergence. As Robinson and Hoffmann put it: "This means that the major axes of variation are not the es along which the . . .[taxa]. . . have differentiated".
Further support for scparatingthe two taxa is provided by the fact that not only are frigidus and sandersoni allopatric over most of their ranges, but where they do overlap (in Newfoundland, Labrador, Manitoba) the specimens examined by us are spht not only by the discriminant analysis but also by a colour character the presence of black pile on the 4th abdominal tergum.
However, if sandersoni and frigidus are to be treated as distinct, two objections must be countered. The first is the close similarity in colour pattern between sandersoni and frigidus in those areas where they arc sympatric. We believe this can be explained in terms of Miillerian mimicry. The remarkable colour convergence between sandersoni and vagans in those parts of their ranges where they occur together is at least as striking as that between sandersoni and frigidus at higher latitudes. It is not unreasonable to postulate a common cause for both sets of chromatic similarity, especially since close parallels can be found in colour convergence involving other species, such as B. rufocinctus Cresson
The second objection springs from the ambiguous status of the Cresson lectotype of Bombus couperi. The possibility that sandersoni and frigidus are merely clinal variants is not yet excluded, although this is perhaps unlikely in view of the NT results. Bearing in mind the efficiency of discriminant analysis in finding statistically significant differences between adjacent regions on a continuum, we would like to examine much larger samples of specimens of both forms taken where they occur sympalrically.
It should, in principle, be possible to establish the degree of reproductive isolation between frigidus and .sandersoni by controlled mating experiments using laboratory cultures. During the pastsummer, using modifications of the technique of Plowright and Jay (1966), we successfully reared three laboratory colonies from a sample of 10 sandersoni queens taken in southern Ontario. The comb of one colony is shown in Fig. 3. We could find no differences m colony development or comb-architecture between sandersoni and frigidus. Both these species appear to be distinct from vagans in that the wax envelopes covering their larvae are more granular in texture, but this, and other slight differences, will need to be confirmed using larger sample-sizes
We propose, in conclusion, that sandersoni remain as a taxon of specific rank, pending further investigation of its relationship to the closely alIied/rigidus. As long as sandersoni is considered specifically distinct from frigidus, or at least until the status of couperi is established by additional analysis, more material, or other characters, we Suggest that the prior epithet couperi Cresson be suppressed because of the questionable affinity of the Cresson lectotype of couperi. However, if sandersoni turns out to be merely a chnat variant offrigidus, then the name couperi should be re-applicd as a sub-specific epithet. Such a procedure is admittedly not entirely satisfactory, but it seems the best that can be done under present circumstances. For purposes of identification, we propose that the relative length of the malar space be used to distinguish sandersoni from vagans where colour convergence between these taxa is
strong, and that the presence or absence of black pile on the 4th abdominal tergum be used to separate sandersom from frigiclus. However, we recognize that these propr)Sa|s may have to be modified as a result of new information.
THE CANADIAN ENTOMOLOGIST
654
strong, and that the presence or absence of black pile on the 4th abdominal tergum be used to separate sandersom from frigiclus. However, we recognize that these propr)Sa|s may have to be modified as a result of new information.
|
|
|
Identification | |
Extracted from: Laverty T.M., & Harder L.D., (1988). The Bumble Bees of Eastern Canada. Can. Ent. 120: 965-987.
Description. All castes small to medium. Head round to slightly elongate; malar space slightly wider than long in female, barely longer than wide in male. Tongue short. Colour as in Figure 13. In most specimens black interalar spot absent or restricted to a few central black hairs; black interalar band present in some specimens. Some males have yellow pile on lateral posterior margins of T5; T6 and T7 black in some specimens. Can be confused with B. vagans vagans and southern forms of B. frigidus.
|
|
|
Names | |
|
|
| Supported by | |
Updated: 2024-04-26 14:22:53 gmt
|